УДК 563.14:553.98.061.31
РАДИОЛЯРИИ КАК ПОТЕНЦИАЛЬНО ВОЗМОЖНОЕ ИСХОДНОЕ ОРГАНИЧЕСКОЕ ВЕЩЕСТВО НЕФТИ
М.С. Афанасьева, М.В. Михайлова (Апрелевское отделение ВНИГНИ)
Решение проблемы происхождения нефтегазовых УВ имеет принципиальное значение, поскольку от него зависят разгадка природы исходного ОВ нефти и оценка нефтематеринского потенциала в связи с поисками залежей УВ нетрадиционного типа. Очевидно, что для выявления природы нефтематеринского вещества необходимо рассматривать его в комплексе с окружающей средой, учитывая ее эволюцию и выясняя роль факторов, влияющих на изменение первичной органики. "Иными словами, вопросы: из чего возникли нефть и газ и как, каким путем они образовались — неотделимы друг от друга" [1, с.58].
Образование конечных продуктов нефтей начинается в определенных участках земной коры, где есть концентрация живых организмов, в прибрежных, богатых жизнью частях моря, и происходит в геологически ничтожное время при существовании благоприятных условий. Расцвет организмов планктона, одноклеточных водорослей, мелких животных,спор определялся взрывами размножения. Размножение живых существ в огромной степени влияет на количество живого вещества в биосфере. В земной коре оно имеет еще большее значение, так как живое вещество представляет собой очаг "активной" энергии, в частности через него проходит процесс передачи энергии Солнца в глубокие слои планеты — образование нефти [2].
В процессе развития Земли менялись бассейны осадконакопления, происходила эволюция живых организмов и их экосистем. Между тем нефтематеринские породы различных возрастов отличаются повышенным содержанием радиолярий. И подобная тесная связь отчетливо прослеживается в течение всего фанерозоя начиная с ордовика [4]. Но наиболее ярко этот необыкновенный феномен природы проявился в классических нефтематеринских породах доманика (среднефранский подъярус верхнего девона) Тимано-Печорской НГП.
Факт исключительно широкого распространения радиолярий в нефтематеринских породах чрезвычайно интересен и наводит на серьезные размышления. Микропалеонтологический анализ радиолярий из классических доманиковых нефтематеринских пород Тимано-Печорского бассейна позволил непосредственно изучить древние организмы радиолярий, являющиеся потенциальным источником ОВ нефти, а также восстановить специфические условия в морских шельфовых бассейнах осадконакопления, способствовавшие концентрации органики, и выявить общие закономерности формирования этих отложений.
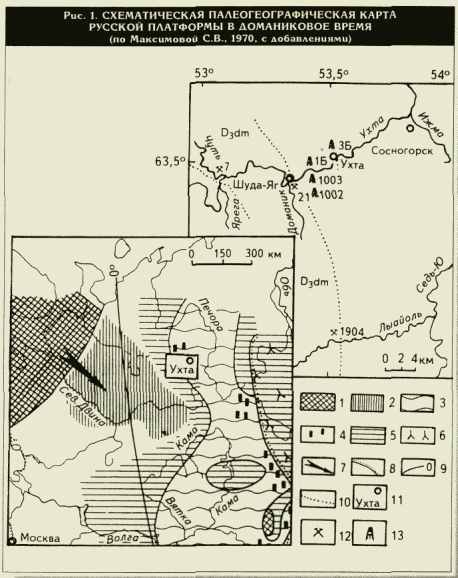
Район исследований расположен на европейском севере России на юге Тимано-Печорской НГП в
окрестностях Ухты (рис. 1). Этот район является стратотипическим для доманика, разрез которого изучен из обнажений по берегам рек Ухта, Доманик, Лыайоль и Чуть (79 литологических образцов и 106 шлифов), а также вскрыт рядом скважин (рис. 2): 1002 Шуда-Яг (18 литологических образцов, 32 шлифа, интервал 66,6-86,5 м), 1003 Шуда-Яг (19 образцов с радиоляриями, 44 литологических образца, 82 шлифа, интервал 68,9-120,0 м), 1Б Ухтинская (14 литологических образцов, 10 шлифов, интервал 67,4-93,6 м) и 3Б Ухтинская (12 образцов с радиоляриями, 42 литологических образца, 46 шлифов, интервал 63,7-118,5 м).Радиолярии — это простейшие одноклеточные организмы. Они относятся к тому же типу царства животных, что и известная всем амеба. Но радиолярии не ползают по субстрату, как амебы: они населяют всю толщу морской воды до глубины 6000 м и не переносят соленость ниже 32 %0, поэтому они обитают только в океанах и морях с нормальной океанической соленостью вод. Радиолярии известны с глубокой древности, с начала развития жизни на Земле, с раннего палеозоя, т.е. почти 500 млн лет. Радиолярии очень малы. Размеры их живой клетки изменяются от нескольких десятков микрометров до 1-3 мм, составляя, как правило, 100-800 мкм. Внутри цитоплазматического живого тела радиолярий находится минеральный скелет, являющийся как бы "балластом", утяжеляющим их клетку. Скелет радиолярий на 98 % состоит из
SiO2 с незначительной примесью Mg, Ca, Al, Na. Внешняя конфигурация скелетов радиолярий поражает своим многообразием. Встречаются шары, икосаэдры, додекаэдры, цилиндры, конусы, эллипсоиды вращения, диски, пропеллеры, треугольники и многие-многие другие причудливые формы, осложненные различными радиальными иглами (рис. 3).Однако как бы ни отличалась одна форма скелета радиолярий от другой, в воде парит лишь шаровидный, эллипсовидный или слегка сплющенный комочек цитоплазмы со своими многочисленными волосовидными аксоподиями (Петрушевская М.Г., 1986). Из цитоплазмы клетки радиолярий наружу могут торчать только кончики самых длинных игл. Скелет у радиолярий внутренний, он не соприкасается с внешней средой, но, вероятно, осуществляет опорную функцию, обусловливающую основную форму цитоплазматического тела радиолярии.
Более того, изучая современные радиолярии, Р.О.Андерсон отметил следующую закономерность: если основной скелет имеет диаметр больше 150-300 мкм, то радиальные скелетные иглы короткие (200-300 мкм), если же диаметр скелета измеряется только несколькими десятками микрометров, то иглы бывают длиной до 1200 мкм. При этом пространство, занимаемое особью, остается примерно одинаковым, тогда как общая удельная масса тел разная
(Anderson R.O., 1983). Предварительные результаты моделирования тела радиолярий показали, что объем цитоплазмы клетки превышает объем кремниевого скелета в 100-700 раз у сферических форм и в 700-3000 раз у иглистых разностей [4] (см. рис. 3).Вероятно, форма скелета, а также соотношение масс скелета и цитоплазмы клетки, скорее всего, определяют глубину погружения радиолярий и выбор оптимальных уровней обитания в толще морской воды. Однако именно существование организма в относительно глубоких водах, более богатых растворенной кремнекислотой, массивного скелета (см.
3, 6, 8 на рис. 3), в то время как приповерхностные виды отличаются развитием более ажурных скелетов (см. 1, 4, 7 на рис. 3). В любом случае организм с большей удельной массой тела будет обитать в более глубоких водах, а не затрачивать неоправданно большое количество энергии на подъем в верхние слои воды.Следовательно, морфологические особенности раковин радиолярий и характер их палеобиоценозов теснейшим образом связаны со средой и изменяются при переходе от приповерхностных к относительно глубоководным, особенно придонным, условиям обитания. Правда, в настоящее время среди геологов устойчиво бытует заблуждение об исключительно пелагическом, планктонном образе жизни всех современных и древних радиолярий и об исключительно глубоководном формировании всех радиоляриевых осадков. Действительно, это верно практически для всех кайнозойских и многих мезозойских отложений (Афанасьева М.С., Вишневская В.С., 1993). Однако при рассмотрении ископаемых радиолярий палеозоя А.В. Хабаков еще в
1932 г. отмечал, что, вопреки распространенному предрассудку, наличие большого количества остатков радиолярий в ископаемом, особенно в древнем, осадке само по себе не может считаться доказательством его абиссального происхождения. Ископаемые радиоляриевые осадки на континентальных щитах относятся в большинстве к батиальным, неритическим, даже лагунным отложениям. Позднее, в 1951 г., О.К. Каптаренко-Черноусова получила фактические данные о незначительных глубинах распространения радиолярий из надмергельных глин киевского горизонта палеогена, а в 1955 г. Е.В. Быкова на материале доманиковых радиолярий доказала, что они могут встречаться в осадках явно прибрежных. Более того, М.Г. Петрушевская еще в 1977 г. впервые предложила гипотезу о происхождении радиолярии от бентосных форм сидячих саркодовых. Между тем все эти факты старательно замалчиваются большинством исследователей, а сам вопрос мелководного и в особенности придонного обитания древних радиолярий усердно обходится стороной. Однако именно эти отличительные черты экологии древних радиолярий имеют решающее значение при определении их роли в формировании УВ-потенциала нефтематеринских пород.Распространение живого вещества на земной поверхности является одним из самых удивительных и исключительных его свойств. И как раз наиболее мелкие организмы размножаются и распространяются с удивительной быстротой. Еще в начале
XIX в. К.Эренберг доказал, что одна диатомея, разделяясь на части, может, если не встретит к тому препятствий, в восемь дней дать биомассу, равную объему нашей планеты, а в течение следующего часа может удвоить это количество органики. Мелкая обычная инфузория-туфелька (Paramaecium) в течение 5 лет может дать массу протоплазмы, объем которой был бы в 104 раза больше объема Земли. Однако это только гипотетические примеры быстроты создания вещества геохимической энергией живых организмов, невероятного вихря миграций элементов, которые они могут вызвать [2].Тем не менее временами действительно удается наблюдать внезапные взрывы жизни в фантастическом размахе. Так, в Адриатическом море иногда отмечается явление, которое итальянцы называют "
mare sporco": огромное внезапное размножение диатомей. Эти одноклеточные водоросли воспроизводятся с такой быстротой, что не хватает времени для восстановления равновесия кремния, растворенного в морской воде. Образуются скопления диатомей, бедных кремнеземом, которые покрывают поверхность моря слоем в несколько метров толщины. Через несколько дней такого размножения диатомовые водоросли оседают на дно в виде студнеобразных масс, быстро поедаемых или разлагающихся. Нарушенное равновесие природы вскоре вновь восстанавливается. Эти примеры дают понятие о потенциальной возможности изменения геохимических процессов, заложенной во всех организмах и радиоляриях в частности. Реально она проявляется в окружающей природе в меньшей степени, не достигая предела, однако порядок явлений от этого резко не меняется [2].Цитоплазма клетки живых радиолярий наряду с различными органеллами содержит запасные жировые включения в виде шарообразных капель размером 3-4 мкм, в редких случаях до 70 мкм. Масса липидов в живой колонии или в клетке
радиолярий достигает 1,5-138,3 мг (Петрушевская М.Г., 1986; Anderson R.O., 1983). Кроме этого, на одну клетку радиолярий приходится до 20-50 симбиотических водорослей размером 2,5-4,5 мкм (Петрушевская М.Г., 1986; Anderson R.O.., Matsuoka A., 1992). Исходя из этого, А.Р.Ормистон [4] сделал предположение о том, что основными поставщиками органики в нефтематеринских породах вместе с водорослями могли быть и радиолярии. Наши исследования и данные геохимии не противоречат этому допущению.Геохимическими исследованиями разных авторов доказана теснейшая связь между белковыми и главным образом липидными компонентами авто- и гетеротрофных организмов и тем битуминозным веществом, которое образуется за их счет сначала в самих осадках, затем в илах, претерпевающих диагенез, и, наконец, в породах, подвергающихся катагенезу [1]. "Соединения, образующие живое вещество — белки, жиры, углеводы, — остаются неприкосновенными и изменяются лишь крайне медленно, не утрачивая своего строения при условии изолированности от явлений жизни" [2, с.172]. Нефть в том или ином виде унаследовала ряд черт, свойственных -именно живому веществу, без которых немыслимо само существование большинства организмов. Живые тела обладают ярко выраженной особенностью по сравнению со всей окружающей неживой материей — диссимметрией, проявляющейся в левизне и правизне пространства жизни. Умирая, в остатках своего вещества, в соединениях своего тела живые организмы геологически долгое время сохраняют диссимметрические структуры, которые отсутствуют в других формах на нашей планете. В земной коре только жизнь и материальные продукты ее разрушения могут обладать диссимметрией, т.е. возможностью проявлений правизны и левизны, их неравенством. "Нефти — тела, обладающие оптически деятельным строением с резким различием правого и левого антиподов. Такого характера соединения углерода создаются только в биосфере и только живым веществом" [2, с.193].
Благодаря этому элементарный состав нефти и его сходство с составом живого вещества являются одной из главных улик биогенной природы исходного вещества нефти [1], а предположение о том, что радиолярии в значительной степени способствуют увеличению органики в осадках, является достаточно вероятным (Афанасьева М.С., Бурзин М.Б., Михайлова М.В. и др., 1995; [3-4]). Массовые скопления радиолярий (вероятно, аналогичные "
mare sporco" диатомей), действительно, могли реально обогащать осадки бассейна сингенетичным ОВ. Последнее явилось результатом посмертной полимеризации цитоплазмы их клеток и впоследствии могло стать одним из источников нефтегазовых УВ доманикоидных пород. Таким образом, устанавливается связь между обилием радиолярий в отложениях эпиконтинентальных морей доманикового типа и высоким качеством нефтематеринских пород, что делает радиолярии хорошим биоиндикатором последних. Для образования нефтематеринских пород, классическим примером которых являются доманиковые отложения Тимано-Печорской НГП, важны не только благоприятные условия среды, обеспечивающие продукцию большого количества органики, но и последующие факторы формирования осадка. С точки зрения нефтяной геологии наибольший интерес представляют шельфовые зоны океанов, окраинные (типа Мексиканского залива) и внутренние моря и крупные озера (типа Каспия), так как осадки именно этих водоемов играют решающую роль в формировании нефтегазоносных бассейнов [1]. В то же время оптимальный вариант для формирования нефтематеринских пород — накопление осадка в условиях бескислородной среды, способствующих сохранению ОВ (Архангельский А.Д., 1927). Всем этим условиям отвечал Тимано-Печорский бассейн в доманиковое время (поздний девон — средний фран).Доманиковые отложения являются нефтематеринскими породами и служат индикаторами особых палеогеографических условий бассейна седиментации. Под термином "доманик" понимается переслаивающаяся
битуминозно-карбонатная толща пород с прослоями горючих сланцев и специфическим комплексом фауны. В настоящее время понятия "доманик" и "доманикит" обозначают не только отложения доманиковой свиты франского яруса, но и разновозрастные породы "доманикового" облика с содержанием Сорг от 2,5 до 10-40 % [1]. Отличительными признаками доманиковых отложений являются: 1) высокое содержание ОВ; 2) характерные литологические фации; 3) специфические комплексы планктонных организмов; 4) отсутствие нормально-морского бентоса.В качестве литологического примера доманиковых отложений рассматривается разрез скв
. 1003 Шуда-Яг (см. рис. 2). В разрезе доманика отмечается закономерное чередование двух основных разновидностей отложений: с одной стороны, черные битуминозные породы, совершенно лишенные каких-либо организмов, но обогащенные Сорг, а с другой стороны, слои, буквально переполненные различными скелетными остатками. Это могут быть как прослои органогенного известняка, так и "присыпки" (россыпь) организмов или их отпечатков по поверхности кремней, кремнистых сланцев или силицитов, не содержащих фауны, которая, возможно, или не сохранилась в них (?), или внезапно погибла именно в моменты их формирований. При этом часто наблюдаются моноорганогенные образования: лингуловые, брахиоподовые, гониатитовые и наутилоидные, тентакулитовые, радиоляриевые и сферовые, скорее всего, водорослевые — акритарховые (12-15 на рис. 4), дополненные массовыми скоплениями зеленых одноклеточных водорослей — тасманитесов (Тельнова О.П., 1995). Особенно широко распространены Tentaculita, встречающиеся по всему разрезу практически во всех разновидностях отложений и чаще всего ориентированные согласно слоистости пород (Кушнарева Т.И., 1963). Однако, образуя иногда на поверхности напластования горючих сланцев сплошные "войлочные" покрытия или обильную россыпь, Tentaculita отсутствуют в толще самой породы. И наоборот, в известняках, как правило, наблюдается хаотичное скопление скелетов Tentaculita именно внутри слоев, где их содержание достигает 85-95 %. Вместе с тем тентакулитовые слои имеют незначительную мощность: не более 5-10 см, а чаще всего — несколько миллиметров. Все это свидетельствует о периодической смене условий, возможно сезонных, в бассейне седиментации и резко различных скоростях выпадения биохемогенных и органогенных осадков.Существует предположение, что в той области, где располагалось Тимано-Печорское море (см.
рис. 1), климат был теплым, тропическим, а палеомагнитный экватор во франском веке проходил через восточную часть Русской платформы примерно по меридиану 50° (Тихий В.Н., 1975). Эта гипотеза, высказанная Д.В.Наливкиным (1956), основывается на присутствии барьерных рифов в Уральском палеобассейне. Развитие рифов свидетельствует о сильном прибое, чистоте воды и тропическом климате со средней годовой температурой до 20 °С. Как подтверждение стабилизации тропического климата можно рассматривать фантастическое разнообразие радиолярий в разрезе доманика (см. рис. 3).Интересным и имеющим принципиальное значение для оценки палеоэкологической обстановки доманикового моря является присутствие в осадках спикул губок: хорошо известно, что все остатки палеозойских губок приурочены только к мелководным отложениям. Однако ранее спикулы губок или только упоминались для отложений доманика (Кушнарева Т.И.,
1959; 1963; Афанасьева М.С., Михайлова М.В., 1995; Афанасьева М.С., Бурзин М.Б., Михайлова М.В. и др., 1995; [3]), или же отрицалась сама возможность развития губок в Тимано-Печорском доманиковом море (Максимова С.В., 1970). Авторами впервые доказано присутствие в осадках доманикового моря кремниевых трехлучевых (стеклянных) и четырехлучевых спикул губок, характеризующихся специфическим нарастанием скелета и наличием центрального канала (см. 2-5 на рис. 4). Впервые обнаруженные в осадках доманика следы ризоидов (см. 11 на рис. 4) также могут свидетельствовать об относительной мелководности доманикового Тимано-Печорского моря, с глубиной 100-200 м даже в наиболее погруженной Ухтинской зоне внешнего шельфа.Перечисленные факты подчеркивают наличие благоприятной палеобиогеографической обстановки накопления потенциально нефтематеринских осадков доманика в тропическом поясе относительно мелководного Тимано-Печорского моря.
В связи с этим намечается ответ на вопрос о причинах чередования в доманиковых и доманикоидных отложениях, с одной стороны, слоев известняков, буквально переполненных скелетами различных организмов, но с низким содержанием Сорг, а с другой стороны, кремнистых разностей, обогащенных ОВ, но практически лишенных
каких-либо скелетных остатков, даже устойчивых к разрушению радиолярий и кремниевых спикул губок.Периодические выбросы больших масс питательных веществ в тепловодных условиях тропического моря способствовали, по-видимому, расцвету фито- и зоопланктона и бентосных организмов, которые и определяли "вспышки" общей биопродуктивности. Это разнообразные аммоноидеи, многочисленные тентакулиты, остракоды из группы энтомозоид, бухиолы, чешуйки рыб и лингулы. Изредка встречаются конодонты и гастроподы, но главное — организмы с кремниевым скелетом (радиолярии и реже губки) и одноклеточные водоросли (
Acritarcha и Tasmanaceae): радиоляриевоводорослевые биоценозы.После естественной биологической гибели организмов осевшее на дно ОВ быстро окисляется кислородом придонного слоя воды за счет жизнедеятельности аэробных бактерий. И если объем ОВ мал, оно разлагается полностью. В этом случае действует сложное сообщество бентосных организмов. Окисляясь кислородом воды до серы, сероводород выходит из реакции, не вызывая отравления воды.
Если же ОВ много, т.е. оно превалирует над минеральной составляющей осадка, то сероводород, не успевая окисляться, отравляет придонный слой застойной необновляемой воды и граница между окислительной и восстановительной средами проходит высоко над осадком в толще воды. Вероятно, именно массовое цветение фитопланктона (возможно, акритарх и тасманитесов) этих стоячих вод, с одной стороны, поставляло огромное количество углеводных веществ биомассы морских водорослей и вызывало невероятный расцвет животной водной жизни, особенно радиолярий. Организмы умирают, не будучи употребленными в пищу другими, их остатки собираются на дне, образуя радиоляриево-водорослевые тафоценозы, а это, с другой стороны, ограничивает аэрацию более глубоких водных слоев, что и вызывает в отдельные моменты гибель бентосных форм и подавление жизнедеятельности аэробных бактерий. Тогда широкое развитие получают анаэробные организмы, главным образом бактерии и некоторые простейшие, обладающие способностью восстанавливать сульфаты и за их счет получать кислород для дыхания.
Фактор сероводородного заражения придонных вод создает совершенно особые условия с господством строго восстановительных реакций с самого начала фоссилизации ОВ. При этом образование сероводородного отравления придонных зон бассейна возможно только в нормальноморской среде, поскольку кислород для дыхания анаэробные бактерии получают при разложении сульфата с выделением столь широко распространенного пирита. Об этом явлении может свидетельствовать пиритизация отдельных прослоев доманиковых пород, а также спикул губок, тентакулитов (см. /,
2 на рис. 4) и скелетов радиолярий: тончайшая решетка их скелетов оказалась состоящей из пирита, заместившего кремнезем (см. 6-10 на рис. 4).Осадки, сформировавшиеся в этих условиях, характеризуются огромным количеством ОВ с широким участием углеводов морских водорослей (акритархи и тасманитесы). Это сапропели, богатые маслянистыми и азотистыми телами, неразрывно связанные с нефтью. И чем больше содержание ОВ в осадке, тем хуже раскристаллизована кремнистая масса породы(Хворова И.В., Дмитрик А.Л., 1972). При этом скелетные остатки организмов просто погружаются в гель и, постепенно растворяясь, сливаются с ним. В битуминозных сланцах часто можно видеть "тени" скелетов разнообразных организмов: раковины их растворились в окружающих кремнях и "слились" с породой при деструкции скелета. Вместе с тем отмечается широкое развитие кремниевой крустификации живой клетки акритарх, сохранившей прижизненную форму и детали скульптуры органической стенки, что в свою очередь указывает на быстрое захоронение организмов в осадках доманика, перенасыщенных кремнеземом (см.
12, 13 на рис. 4).Тем не менее благоприятная экологическая ситуация постепенно восстанавливается: вновь набирает силу расцвет жизненных форм, вновь происходит массовое цветение фитопланктона и вновь все это заканчивается сероводородным заражением и гибелью организмов. Такие "волны жизни" исчезают так же быстро, как и появляются [2]. Итак, мы наблюдаем закономерную циклическую смену радиоляриево-водорослевых тафоценозов, отражающих различные условия существования организмов в среднефранском доманиковом море.
Во время пиков кислородной недостаточности в придонных водах эпиконтинентальных морей настоящего и прошлого радиолярии вытесняют из биоценозов нормально-морские бентосные и планктонные организмы: фораминиферы, остракоды, гониатиты, тентакулиты, губки — и ассоциируются только с акритархами. Более того, фораминиферы вообще нехарактерны ни для доманиковых, ни для доманикоидных отложений (Быкова Е.В., 1955). Предположение, что фораминиферы существовали в подобном бассейне осадконакопления, но раковины их растворились, маловероятно, поскольку известковые скелеты иных организмов прекрасно сохранились. По-видимому, только важные биологические отличия между этими группами организмов могут объяснить феномен подобного кратковременного, но яркого доминирования радиолярий. Дело в том, что жизненный цикл развития фораминифер включает чередование полового и бесполого поколений. Во время полового размножения гаметы опускаются на морское дно для последующего воспроизводства и развития, которое было бы невозможно при отсутствии или недостатке кислорода. Радиолярии же размножаются путем бесполого деления клетки и не зависят от кислородных условий придонных вод [4]. Следовательно, обилие радиолярий в отложениях эпиконтинентальных морей прошлого может являться признаком кислородного минимума придонных вод.
Недостаток кислорода мог благоприятствовать сохранению нейтральной реакции в водах палеоокеанов и образованию кислых продуктов обмена на клеточном уровне. Эти факторы препятствовали формированию кальцитового и фосфатного скелетов у живых организмов. Но эти же условия палеосреды могли бы быть оптимальными для развития скелетов из опала — геля ортокремниевой кислоты. Кроме того, в процессе эволюционного развития радиолярии изначально стали образовывать свои раковины из сернокислого стронция (
SrSO4): возможно, сульфатные воды палеоморей были более благоприятны для синтеза целестина. Запись об этих событиях сохранилась в генах у Acantharia и реализовалась в форме целестинового скелета, который они, видимо, смогли пронести через весь фанерозой. Однако в целом основное направление эволюции радиолярий пошло по пути отказа от сернокислого стронция и начала синтеза опала (Si2-nН2О) (Афанасьева М.С., Вишневская В.С, 1992).Эти маленькие нюансы эволюции радиолярий помогут разгадать причины, вызвавшие отсутствие фораминифер (простейших с известковыми раковинами) и способствовавшие расцвету радиолярий (организмов с кремниевым скелетом) в своеобразных экологических условиях бассейнов доманикового типа.
Предложенный нетрадиционный подход к решению вопроса природы ОВ нефти затронул серьезную проблему возможного источника нефтематеринского вещества и показал необходимость дальнейшего основательного обсуждения важной роли радиолярий в формировании УВ-потенциала нефтематеринских пород.
Выводы
1. Образование нефтематеринских доманиковых отложений могло иметь место только при определенной фациальной обстановке продуцирования и консервации исходного ОВ нефти:
скопление ОВ с преобладанием жировых включений клеток радиолярий и с примесью значительного количества растительных углеводов (акритархи и тасманитесы);
отложение исходного материала в восстановительной среде нормально-морского бассейна с отравленным сероводородом придонным слоем;
теплый и влажный климат, обусловливающий быстрое воспроизводство и накопление больших масс ОВ.
2. Обилие радиолярий в отложениях эпиконтинентальных морей прошлого, вероятно, является признаком кислородного минимума придонных вод.
3. Радиолярии не только обогащали осадки доманйкового моря биогенным кремнеземом, но и, скорее всего, были основными поставщиками органики.
4. Предлагается использовать данные о формировании богатых радиоляриево-водорослевых тафоценозов в качестве обоснования при прогнозировании нефтяных месторождений нетрадиционного типа.
Литература
1. Вассоевич Н.Б. Геохимия органического вещества и происхождение нефти. — М.: Наука, 1986.
2. Вернадский В.И. Очерки геохимии. — М.: Наука, 1983.
3. Вишневская
B.C., Мерц А.В., Седаева К.М. Девонские радиолярии и их роль в образовании нефти // Докл. РАН. — 1993. — Т.ЗЗЗ, № 6. — С.745-749.4. Ormiston A.R. The association of radiolarians with hydrocarbon source rocks // Micropaleontology. Special publication.
— 1992. — № 6. — P.9-16.©М.С.Афанасьева, М.В.Михайлова,
1998
To solve a problem of oil and gas hydrocarbons source origin has a principal significance in oil and gas geology because it would allow to understand the nature of oil's original organic matter and estimate oilsour-ce potential of rocks. In the process of the Earth's evolution the sedimentary basins have been changed, and evolution of live organisms and their ecosystems took place. Meanwhile, oil source rocks of different ages are characterized by increased content of Radiolaria. An similar close relationship is distinctly traced during the whole Pha-nerozoic from Ordovician period. Using of micropaleontologi-cal analysis with reference to Radiolaria from classic domanic oil source rocks of Timano-Pec-hora basin allowed directly to study ancient organisms of Radiolaria which are potential source of oil organic matter as well as to reconstruct the specific conditions in marine offshore sedimentary basins contributed to organics concentration, and to recognize general regularities of these deposits formation. The role of Radiolaria in forming hydrocarbon potential of oil source rocks is discussed. Abundant Radiolaria in deposits of ancient epicontinental seas appear to be an indication of oxygen minimum of near-bottom waters. Data on forming rich radiolarian-algal taphocoenosis are proposed to be used as a basis for forecasting non-traditional oil fields.
1 —
суша; 2 — области отсутствия отложений вследствие последующего размыва; 3 — доманиковые отложения; 4 — кремниевые линзы и пропла-стки в доманиковых отложениях; 5 — морские отложения недоманикового типа; 6 — зона подводного вулканизма; 7 — направление сноса обломочного материала; 8 — граница платформы; 9 — палеомагнитные широты, град; 10 — геологическая граница;11 — район работ; 12 — обнажения; 13 — скважины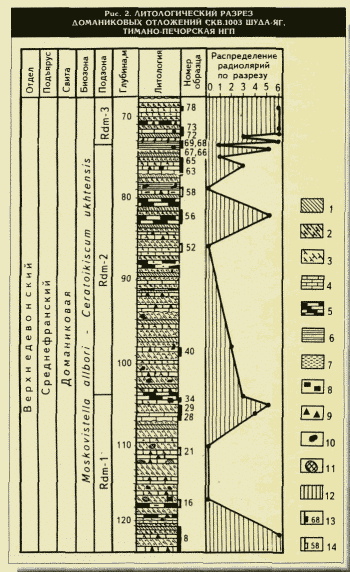
1 —
кремень; 2 — кремнистый сланец; 3 — окремнение; 4 — известняк; 5 — кремнеизвестняк; 6 — глина; 7 — глинистость; 8 — пирит; 9 — битуми-нозность; 10 — карбонатные конкреиии; 11 — кремниевые конкреиии; 12 — Radiolaria; 13 — номер образца с радиоляриями; 14 — номер образца без радиолярийРаспределение радиолярий по разрезу в баллах, экз/г породы:
1 — единично (1-5), 2 — редко (10-15), 3 — нередко (20-40), 4 — часто (50-100), 5 - очень часто (200-500), б - в массе (>1000)

1 — Palaeothalomnus arrhinia (Foreman):
ФФ-П001/085-04803; штрих =67 мкм; 2 — Palaeothalomnus quadriramosum Foreman: ФФ-П001/105-11701; штрих =56 мкм; 3 — Palaeothalomnus timokhinl Afanasieva: ФФ-П001/086-11620; штрих=50 мкм; 4 — Borisella maksimovae Afanasieva: ФФ-П001/036-04919; штрих=83 мкм; 5 — Moskouistella khaini Afanasieva: ФФ-П001/041-04727; штрих=67 мкм; б — Spongentactinella veles (F о r е m a n): ФФ-П001/065-04918; штрих=67 мкм; 7 — Astroentactinia vishnevskayae Afanasieva: ФФ-П001/034-08907; штрих=5б мкм; 8 — Entactlnosphaera menneri Afanasieva: ФФ-П001/014-04714; штрих=50 мкм. Скв.1003 Шуда-Яг: 1, 5, 8 — образец 68 (интервал 73,0-73,3 м); 2, 3 — образец 73 (интервал 71,4-71,9 м); 4,6 — образец 78 (интервал 68,9-69,3 м); скв. 3Б Ухтинская: 7 — образец 116 (интервал 100,6-101,0 м)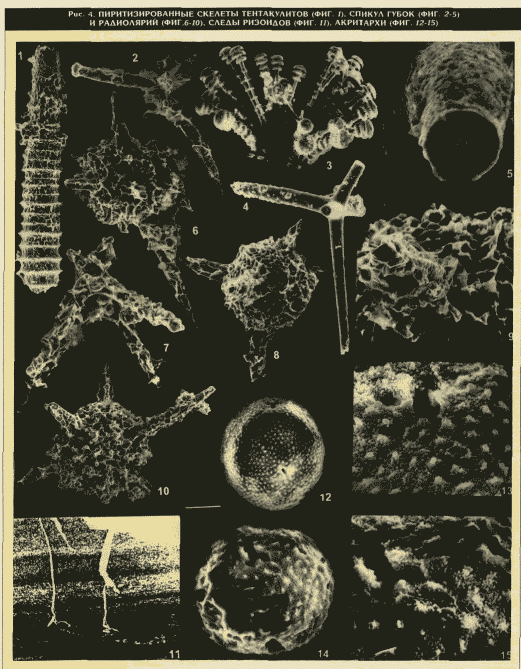
1 — Tentaculites semilukianus G. Ljaschenko:
ФФ-П004/001-09028, мapкep=100 мкм; 2 — Tetraxonida (?) sp.3: ФФ-П003/004-09101, мapкep=42мкм; 3 - Triaxonida (?) sp.l, ФФ-П003/001-04320, мapкep=40мкм; 4, 5 - Triaxonida (?) sp.4, ФФ-П003/004-09817: 4 — мapкep=10 мкм, 5 — мapкep=67 мкм; 6 — Entactinosphaera cf. menneri Afanasieva; ФФ-П001/014-09133; мapкep=42 мкм; 7 — Palaeoscenidium cf. ctadophorum D e f l a n d r e: ФФ-П001/078-09135; мapкep=33 мкм; 8,9 — Ornatoentactinia sp.: ФФ-П001/044-09123; 8 — мapкep = 45 мкм, 9 — мapкep = 14 мкм; 10 — Entactinosphaera cf. grandis N a z a r o v: ФФ-П001/016-09119; мapкep=77 мкм; 11 — следы ризоидов, шлиф: мapкep=3 мм; 12-15 — Acritarcha (?) gen. et sp. indet.: 12, 13 — ФФ-П002/001-04603: 12 — мapкep = 50 мкм, 13 - мapкep =10 мкм; 14, 15 — ФФ-П002/001-08924: 14 — мapкep=23 мкм, 15 — мapкep=10 мкмckb.
3б Ухтинская: 1, 2, 6, 7-10 — образец 114 (интервал 104,2-104,7 м); скв. 1003 Шуда-Яг: 3 — образец 29 (интервал 105,0-106,0 м); 11 — образец 21 (интервал 110,2-111,0 м); 12, 13 - образец 34 (интервал 104,1-104,6 м); 13-15 — образец 40 (ин-тервал 98,0-99,0 м); p. Лыайоль, обнажение 1904: 4, 5 — образец 6